
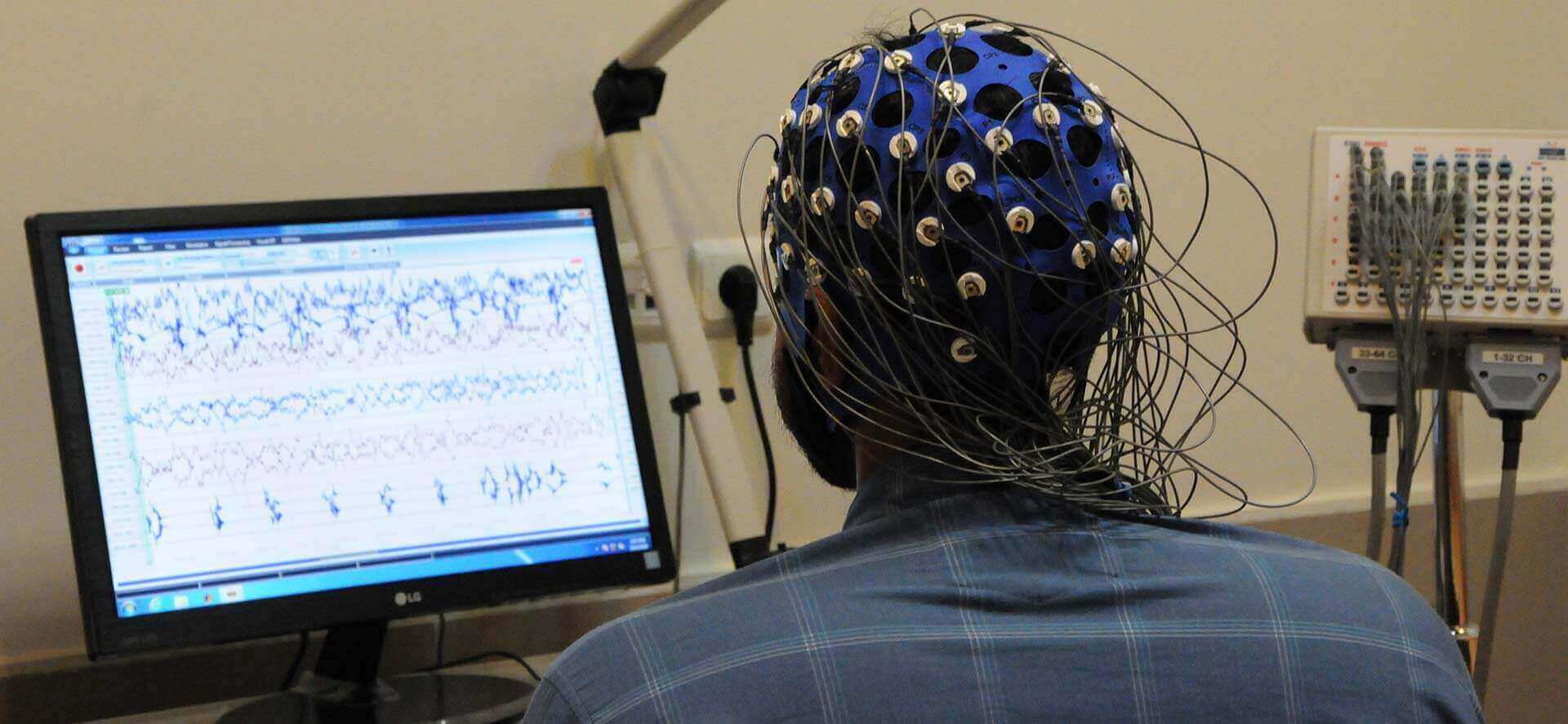





دهمین همایش بینالمللی علوم شناختی بعد از ظهر روز جمعه با تقدیر از صدها نفر از استادان، پژوهشگران، دانشجویان و متخصصان شرکتکننده در نشستها و برنامههای کوناگون و نیز با معرفی بهترین پوسترهای علمی ارائه شده به همایش به کار سه روزه خود پایان داد. دهمین همایش روز چهارشنبه ۲۶ اردیبهشت ۱۴۰۳ با سخنان دکتر روحالله دهقانی فیروزآبادی معاون علمی، فناوری و اقتصاد دانشبنیان رییس جمهور، دکتر سید کمال خرازی رییس هیات امنای موسسه آموزش عالی علوم شناختی، و دکتر سهراب حاجیزاده و دکتر مهدی تهرانیدوست، به ترتیب رییس و دبیر علمی همایش، کار خود را آغاز کرد...






